Обзор литературы. Технология возобновления леса в ханты-мансийском автономном округе Как называется подрост деревьев
Текст Бориса Колесова:
(1) Ясное морозное зимнее утро. (2)По узкой деревенской тропке шагаю с ведром на родник. (З)Лет мне пока не так много, чтобы за один раз принести два ведра воды. (4)Это потом уже будут и две объёмистые оцинкованные посудины, и даже коромысло. (5)По примеру бабушки отправлюсь к источнику хорошо вооружённым деревенским вспомогательным устройством, но не смогу балансировать вёдрами — стану ходить по-прежнему.
(6) Вот, значит, невеликим мужичком иду к оврагу, глубокому и заснеженному, где струится чистый, никогда не замерзающий ручей. (7)Вижу наверху, за белым провалом, за небесно синеющими сугробами, зелёные ёлочки. (8)И почему-то радостно на душе становится и хочется вприпрыжку бежать в овраг, чтобы потом, уже на выходе из него, обернуться и снова приметить зелёные живые деревца. (9)Вместе с ними и я доволен прозрачностью небесной, белизной снегов и бодрым, не очень сильным морозцем.
(10)Потом, уже летом, не раз ходил мимо тех ёлочек за три километра в соседнее село.
(11) И всегда встречал их на дороге — пообочь тропки — с такой восторженной радостью, словно ничего не видел на земле более прекрасного! (12)А может, и вправду не видел такого, чтоб вровень с их густой изумрудной прелестью: в нашей деревушке, затерянной по февралю в снегах, а по июлю в черёмуховых овражистых дебрях, не было ни картинных галерей, ни даже клуба.
(13)Сейчас мне уже под семьдесят, но всегда с замиранием сердца вспоминаю те ёлочки.
(14) Не могу объяснить, что со мной, — иногда аж слеза прошибает: дорогие вы мои, ненаглядные!
(15) Дни меж тем продолжались чередой годов; многие события, встречи становились привычными, потихоньку стирались воспоминания. (16)Но никуда не ушли те дни, когда водили ноги досужего зрителя по заповедной целине Приокско-террасных ельников, по дивному тамошнему заповеднику. (17)Ели тут были особенными. (18)Сам заповедник, со всеми его растениями и животными, необычайно интересен. (19)Одни зубры чего стоят! (20)Где ещё увидишь могучих великанов, существовавших в пору древнеславянских народов? (21)Но что касается заповедного елового подроста… (22)Тут держишь ухо востро!
(23) По ту сторону Оки, на лесостепном или степном юге, трудновато уже встретить ёлочку, произрастающую вполне естественно. (24)В Средней полосе России для подобных деревьев природой положен предел, проведена невидимая черта, объясняемая особенностями российского климата. (25)Взять хоть влагу, до которой ели весьма охочи, — маловато её в степях* разве не так? (26)В климате более сухом и жарком выстоять маленькой ёлочке, быстро подрасти, чтобы поглубже в землю пустить корни и набрать силу, не так-то легко. (27)Даже искусственные лесонасаждения, где подросту обеспечен заботливый уход, получаются, говоря языком лесоводов, нерентабельными, и сосны имеют предпочтение среди хвойных пород. (28)Аккуратные зелёные строчки молодых сосен я встречал даже на степном Дону. (29)А вот еловых посадок не видел.
(30)Высоко ценится ель в деревообработке, в бумажном производстве, в изготовлении певучих музыкальных инструментов… (31)Но как оценить красоту её чудесного зелёного убранства посреди русских снегов?
(32) Природа наша столь удивительна, что возникает желание быть с ней заодно в стрем-лении жить, побеждать трудности и быть полезным людям. (ЗЗ)Бывали у меня радостные, счастливые дни. (34)Бывали и такие, когда деловой настрой заставлял понимать, что важно в повседневном, полном всяческих хлопот, быту людей. (35)Кому книжку подавай, кому скрипку, кому дрова для печки, кому плахи какие-нибудь для строительства дома.. (Зб)Кто-кто, а лесоводы знают, для чего пестуют искусственные посадки.
(37) Но болит и болит сердце за еловый лес… (38)3а те ельни, как называли в старину влажные лесные места с преобладанием этой породы хвойных, за те посадки в заказниках, где молодые ёлочки подвергаются беспощадному наступу. (39)Кто на них, бедных, наступает в наши времена? (40)3ачастую дикие заготовители стремятся срубить дерево крупное и сильное, чтобы взять от него пышную верхушку. (41)Кроме того, в больших посадках, где способ ную охрану не поставишь, полным ходом идёт заготовка дров. (42)Может, найдутся такие, что станут мне возражать, но откуда тогда в молодых лесах столько безобразных пней, а?
(43) Берегите чудо природы, берегите красавиц, особенно в предновогодние дни.
(44) Молодые ёлочки — они ведь для всего и для всех. (45)Молодёжь для страны — золотой фонд. (46)3елёные красавицы заказников — благо для русского леса. (47)Нежные эти ёлочки — молодёжь вполне достойная.
(По Б. Колесову)
Борис Колесов — российский писатель, журналист, сценарист.
Сочинение по тексту:
Что является истинной красотой природы? Над этим вопросом рассуждает в тексте российский писатель, журналист Борис Колесов.
Автор вспоминает свое детство, как он, направляясь к источнику за водой, встретил на своем пути зеленые ёлочки. Казалось бы, обычные ели, каких много повсюду, но Б.Колесов запомнил их на всю свою жизнь. Спустя много лет автор не может забыть эмоций, которые ему доставляли "зеленые красавицы".
Позиция автора ясна: природа красива. Мы должны уметь видеть эту красоту и беречь ее.
Я разделяю мнение Бориса Колесова. Действительно, нужно уметь видеть и ценить красоту, которая окружает нас. Ведь природа не прячет ее от нас.
Обратимся к стихотворению Александра Сергеевича Пушкина "Зимнее утро". Лирический герой восхищен красотой зимнего утра. Автор всячески оживляет картину, заставляя читателя проникнуть в пейзаж прекрасного зимнего утра и прочувствовать все самому. Чтобы показать всю красоту, автор обращается к такому средству художественной выразительности, как олицетворение: "вьюга злилась", "мгла носилась".
В рассказе Василия Шукшина "Старик, солнце и девушка" восьмидесятилетний старик, будучи слепым, каждый день в одном и том же месте любовался природой. Рассказ заставляет задуматься над тем, что человек не только видит красоту природы, но и чувствует её.
Борис Колесов уверен, намного лучше жить в единстве с природой, чем врозь. Такие эмоции, которые доставляет человеку природа и ее красота, нигде не найти.
Это слово - "кукольник", что объясняется довольно просто. Все, что связано со словом "кукла," ассоциируется с чем-то маленьким, связанным с подрастающим поколением, вот и для "деток" подобралось слово.
Небольшая справка о "подросте":
Само по себе слово "подрост" обозначает поколение молодых деревьев, которое выросло либо в самом лесу под пологом более старых деревьев, либо на пустом месте - это могут быть вырубленные или выгоревшие участки.
По возрастному признаку деревья из подроста относятся к молодняку.
Практическое значение "подроста" довольно значительно: именно участки с молодыми деревьями могут стать основой нового лесного массива.
Люди давно поняли важность таких "подростов" для сохранения лесов. Поэтому помимо естественных участков с молодыми деревьями можно встретить и искусственные, то есть специально подсаженные, чаще встречаются комбинированные. Специалисты оценивают качественные показатели, породы, густоту имеющихся естественных подростов по показателю количества деревьев на определенную единицу площади и подсаживают новые экземпляры, доводя показатели густоты насаждений до установленной оптимальной нормы и, тем самым закладывая основу для новых ярусов леса.
Помимо контроля за подростом специалисты лесного хозяйства применяют ряд практических мер, способствующих правильному формированию лесного массива, например, различные виды рубки, которые имеют свое назначение и специфику.

Введение. Изучение естественного возобновления имеет особую значимость в лесоразведении. Такие исследования позволяют определить количество и качество молодого поколения, идущего на смену материнского древостоя. Большой интерес вызывает установление устойчивости насаждений с преобладанием сосны обыкновенной.
Текущий прирост молодых деревьев является объективным показателем для оценки состояния подроста, так как древостой может оказывать на него как отрицательное, так и положительное воздействие .
Известно, что количество и качество подроста под пологом леса во многом зависит от породы, слагающей древостой. На преимущество естественного возобновления с точки зрения биологии и экономики подчеркивал Г.Ф. Морозов .
Вайс А.А. установил, что в процессе роста в насаждении возрастает влияние деревьев друг на друга .
Итешина Н.М., Данилова Л.Н., Петров Л.В. определили, что сосновые древостои естественного происхождения имеют более низкие темпы роста по сравнению с искусственными насаждениями .
Основными источниками, раскрывающими вопросы морфологических взаимосвязей которые влияют как на рост, так и на состояние насаждений явились работы Вайса А.А. .
Целью исследования являлась оценка роста молодняка в высоту под пологом и на открытом месте.
Объекты и методика исследований. Объекты исследований располагались на территории Усинского лесничества. Для проведения исследований подбирались участки леса с полнотой 0,4, 0,7 и открытое место.
Работы проводились в конце вегетационного периода – в сентябре. Всего было заложено 3 пробной площади. На каждой пробной площади размещалось 30 опытных площадок 1*1 м, где проводился выборочный перечет подроста сосны (всходы не учитывались) с измерением линейных показателей. На отобранных пробных площадях проведены измерения приростов, возраста по мутовкам, замер высоты, диаметр кроны и диаметр ствола у подроста сосны обыкновенной в разных условиях освещенности (разнополнотные древостои).
Обмерены годичные приросты в высоту 131 подроста сосны на 3 пробных площадях .
Экспериментальные исследования. Прирост деревьев в сосняках является важнейшим таксационным показателем как отдельно стоящего дерева, так и всего древостоя. С помощью прироста можно оценить потенциальную продуктивность древостоя, добротность условий местопроизрастаний, конкурентные взаимоотношения между компонентами насаждений.
Текущий прирост молодых деревцев сосны обыкновенной является объективным показателем, который характеризует их рост и состояние, а также синтезирует результаты жизнедеятельности растительного организма.
Для идентификации прироста молодых деревцев в высоту с целью разделения подроста сформированных под влиянием экологических факторов были построены графики зависимости прироста по годам и прироста с учетом возраста. Данные графики приведены ниже.
Рисунок 1. Зависимость прироста молодых деревцев по годам
Проанализировав график зависимости приростов по годам можно сказать, что на открытом месте минимальный прирост подроста наблюдался в 2008 году, а максимальный прирост зафиксирован в 2011 году. Также можно сказать, что средний прирост варьирует в пределах 7 см. Под пологом древостоя минимальный прирост подроста наблюдался в 2000 и 2003 году, максимальный прирост в 2005 году. Прирост подроста сосны по высоте по годам в абсолютном выражении показал его динамичность. Изменение прироста в пределах пробной площади может различаться по всем годам, либо иметь сходство в отдельные годы (рис.1).
Данные изменения в приросте сосны обыкновенной можно объяснить тем, что в разные годы климатические факторы по-разному влияли на рост подроста сосны обыкновенной.
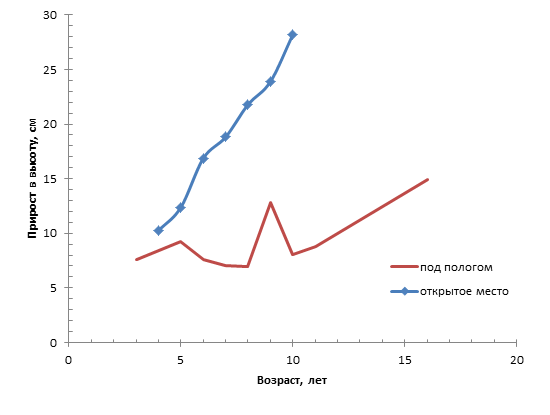
Рисунок 2. Зависимость прироста подроста от возраста
Из графика зависимости прироста с учетом возраста видно, что на открытом месте с увеличением возраста прирост увеличивается (прямая зависимость). Минимальный прирост подроста наблюдался в возрасте 4 лет, максимальный в 10 лет. На открытом месте интенсивность роста тем больше, чем крупнее подрост. Под пологом леса минимальный прирост наблюдался в возрасте 8 лет и резко увеличился в возрасте 9 лет. Под пологом нет строгой закономерности как на открытом месте, так как на прирост под пологом большое влияние оказывают лимитирующие факторы (свет, биогенные вещества, течение и давление среды, почва, пожары, микросреда и т. д.).
В состоянии самосева молодые деревца имеют малоразвитую корневую систему и небольшую листовую поверхность. По мере разрастания корней и листьев ассимилирующая способность крон возрастает, увеличивается прирост древесины. Плохой рост подроста сосны, под пологом взрослых древостоев, обусловлен не только недостатком света, но и тем, что деревья материнского древостоя своими мощными и хорошо развитыми корнями перехватывают питательные вещества и влагу из почвы. На жизнедеятельность корней существенно влияет вода, находящаяся на иллювиальном горизонте, которая чаще всего возникает весной после таяния снега. При высоком ее стоянии задерживается жизнедеятельность корней, снижается продолжительность вегетационного периода и, как следствие, уменьшается прирост. Сосна – светолюбивая порода и хорошо растет только без затенения. Следовательно, на открытом месте интенсивность прироста значительно выше, чем под пологом.
Вывод. Изучив распределение молодняка по возрастам и по годам на открытом месте и под пологом можно сделать следующие выводы:
На открытом месте минимальный прирост подроста наблюдался в 2008 году, а максимальный прирост выявлен в 2011 году;
Под пологом древостоя минимальный прирост подроста наблюдался в 2000 и 2003 году, максимальный прирост в 2005 году;
Величина прироста в пределах пробной площади может различаться по всем годам, или иметь сходство в отдельные годы;
На открытом месте интенсивность роста тем больше, чем крупнее подрост;
Под пологом нет определенной закономерности как на открытом месте, так как на прирост под пологом большое влияние оказывают лимитирующие факторы (свет, биогенные вещества, течение и давление среды, почва, пожары, микросреда и т. д.);
На открытом месте интенсивность прироста значительно выше, чем под пологом.
Библиографический список
- Ангальт Е.М., Жамурина Н.А. Анализ прироста соснового подроста и молодняка в сосново-ясеневых культурах [Текст] //Известия Оренбургского государственного аграрного университета. 2013. № 6 (44).С 31-34.
- Морозов Г. Ф. Избранные труды / Москва: Лесная промышленность, 1971. 536 с.
- Вайс А.А. Динамика конкурентных взаимоотношений: в в- между древесными особями в сосновом ценозе [Текст] // Вестник КрасГАУ. 2011. № 5. С. 84-87
- Итешина Н.М., Данилова Л.Н., Петров Л.В. Рост и строение сосновых древостоев в различных лесорастительных условиях [Текст] // Инновационному развитию АПК и аграрному образованию – научное обеспечение: Материалы Всероссийской научно-практической конференции. 2011. Т.1. С. 265-267
- Вайс А.А. Динамика конкурентных взаимоотношений между древесными особями в сосновом ценозе [Текст] // Вестник Красноярского Государственного аграрного университета. 2011. №5. С.84-87.
- Вайс А.А. Связь диаметров нижней части стволов сосны обыкновенной (Pinus sylvestris L.) в условиях Средней Сибири [Текст] // Лесная таксация и лесоустройство. 2011. № 1-2. С.29-32.
- Вайс А.А. Закономерности связи диаметров деревьев лиственницы сибирской (Larix sibirica) на высоте груди и диаметров на высоте пня в условиях средней и южной сибири [Текст] // Вестник Адыгейского Государственного университета. Серия 4: Естественно- математические и технические науки. 2011. № 1. С. 53-60.
- Мачык М.Ш., Вайс А.А. Оценка хода роста морфологических параметров молодняков сосны с учетом их возрастной структуры в условиях Восточнотувинско-Южнозабайкальской горной лесорастительной области [Текст] // Современные научные исследования и инновации. 2016. № 1 (57). С 268-276.
Оценка жизненного состояния хвойного подроста
К категории методических пособий можно также отнести книги, написанные и изданные нашими друзьями и коллегами, и продающиеся в нашем некоммерческом Интернет-магазине :
| Организация
исследовательской и проектной деятельности в
школе
(пособие для учителя) Организация образовательного процесса в массовой школе на основе проектной и исследовательской деятельности , которая коренным образом изменяет функцию учителя, наталкивается на неподготовленность педагогических кадров к претворению инновационной идеи. Снова учитель один на один оставлен с серьёзными проблемами, возникающими при освоении инноваций. Представляемое пособие для учителя призвано объяснить введение в школьный образовательный процесс таких инновационных технологий как исследовательская и проектная деятельность . |
|
| Основы гидроботанических исследований: методическое
пособие-определитель
Учебно-методическое пособие посвящено теоретическим и практическим вопросам, связанным с организацией гидроботанических исследований учащихся. Даны рекомендации к организации (подготовке и проведению) гидроботанических исследований и конкретные методики , относящиеся к определённым направлениям гидроботанических исследований. Методики, разработанные ведущими учёными в области гидроботаники, адаптированы для восприятия и понимания и предполагают свободное овладение ими самостоятельно или под руководством педагогов. Приводятся ключи для определения растений водной флоры в полевых условиях, справочные материалы по некоторым наиболее важным направлениям гидроботаники. |
|
 |
От наблюдения до выступления. Пособие для педагога
В пособии рассказывается, как вы можете оказаться на природе, найти там интересные для вас объекты исследования, понаблюдать за ними, измерить, сравнить, сфотографировать, оформить собранный материал в виде доклада, исследовательской работы и выступить на конференции. Пособие может быть полезно юным натуралистам, которым интересно знакомиться с природой и раскрывать ее тайны, учителям, работающим со школьниками по подготовке к конференциям. Пособие поможет заинтересовать и организовать ребят, сформировать у них множество полезных и нужных УУД (универсальных учебных действий) по ФГОС. |
Данное пособие может пригодиться руководителям детских туристских кружков, а также будет интересно широкому кругу любителей самодеятельного туризма, так как содержит сведения по подготовке и проведению не очень сложных путешествий (от походов выходного дня до многодневных пеших или байдарочных). В книге, кроме специальных туристских сведений, описаны также способы приготовления походной еды, первая помощь, игры в свободное время и многое другое, что может пригодиться в туристском походе. В данном пособии содержится много полезных советов, основанных на многолетнем опыте автора, поэтому желательно его внимательно изучить до похода и обязательно взять с собой. |
| Другие методические материалы по
организации проектной деятельности и исследовательской работы школьников в природе Пособие для учителя " |
480 руб. | 150 грн. | 7,5 долл. ", MOUSEOFF, FGCOLOR, "#FFFFCC",BGCOLOR, "#393939");" onMouseOut="return nd();"> Диссертация - 480 руб., доставка 10 минут , круглосуточно, без выходных и праздников
Гуталь Марко Миливоевич. Жизнеспособность и структура подроста ели под пологом древостоев и на вырубках: диссертация... кандидата сельскохозяйственных наук: 06.03.02 / Гуталь Марко Миливоевич;[Место защиты: Санкт-Петербургская государственный лесотехнический университет имени С.М. Кирова http://spbftu.ru/science/sovet/D21222002/dis02/].- Санкт-Петербург, 2015.- 180 с.
Введение
1 Состояние проблемы 9
1.1 Общие сведения о еловых фитоценозах 9
1.2 Подрост ели 11
1.2.1 Особенности возрастной структуры подроста ели 12
1.2.2 Особенности светового режима под пологом ельников 16
1.2.3 Жизнеспособность подроста ели 22
1.2.4 Численность подроста ели 25
1.2.5 Влияние типа леса на подрост ели 27
1.2.6 Особенности развития подроста ели под пологом древостоя 30
1.2.7 Влияние растительности нижних ярусов на подрост ели 33
1.2.8 Влияние хозяйственных мероприятий на подрост ели 35
2 Программа и методика исследований 39
2.1 Программа исследований 39
2.2 Исследование лесного фитоценоза по структурным элементам 40
2.2.1 Определение основных характеристик древостоя 40
2.2.2 Учет подроста 41
2.2.3 Учет подлеска и живого напочвенного покрова 46
2.2.4 Определение биометрических показателей хвои 49
2.3 Объекты исследований 51
2.4 Объем выполненных работ 51
3 Динамика состояния подроста ели под пологом древостоя .
3.1 Динамика жизненного состояния подроста ели по результатам длительных исследований 53
3.2 Закономерности изменения жизнеспособности елового подроста в связи с типом леса 69
3.3 Влияние материнского полога на динамику состояния и структуру подроста ели
3.4 Связь жизнеспособности подроста ели с величиной среднего прироста за период 3, 5 и 10 лет.
3.5 Возрастная структура как индикатор состояния подроста 86
3.6 Структура по высоте подроста как индикатор состояния 89
3.7 Сравнительный анализ состояния и структуры подроста ели в ельниках Лисинского и Карташевского лесничеств 93
4 Влияние хозяйственых мероприятий на численность и жизнеспособность подроста ели
4.1 Влияние рубок ухода на динамику жизнеспособности подроста ели 105
4.2 Изреживание подлеска – как мера содействия естественному возобновлению ели 122
5 Динамика состояния подроста ели на вырубке 127
5.1 Особенности структуры и состояния подроста ели 127
5.2 Зависимость динамики состояния подроста ели от давности рубки 134
6 Биометрические характеристики хвои как индикатор жизнеспособности подроста ели
6.1 Биометрические показатели хвои под пологом и на вырубке 140
6.2 Биометрические показатели хвои жизнеспособного и нежизнеспособного подроста ели.
Список литературы
Особенности светового режима под пологом ельников
Ель – это одна из главных лесообразующих пород на территории РФ, занимающая четвертое место по занимаемой площади, уступая только лиственнице, сосне и березе. Ель произрастает от тундры до лесостепи, но именно в таежной зоне в наибольшей степени проявляется ее лесообразующая и эдификаторная роль. Род ель (Picea Dietr.) относится к семейству сосновых (Pinacea Lindl.). Отдельные представители рода ели ведут свое начало с мелового периода, то есть 100-120 миллионов лет назад, когда у них был один общий ареал на евроазиатском континенте (Правдин, 1975).
Ель европейская или обыкновенная – (Picea abies (L.) Karst.) широко распространена на северо-востоке Европы, где образует сплошные лесные массивы. В Западней Европе хвойные леса не являются зональным типом растительности, и там имеет место вертикальная дифференциация. Северная граница ареала в России совпадает с границей лесов, а южная доходит до чернозмной зоны.
Ель обыкновенная – дерево первой величины с прямым стволом, конусовидной кроной и не строго мутовчатым ветвлением. Максимальная высота достигает 35-40 метров в равнинных условиях, а в горах встречаются экземпляры высотой до 50 м. Самое старое известное дерево имело возраст 468 лет. Однако, возраст более 300 лет очень редок, а в полосе хвойно-широколиственных лесов снижается до 120-150 (180) лет (Казимиров, 1983).
Ель обыкновенная характеризуется относительно большой пластичностью корневой системы, способной приспосабливаться к различным почвенным условиям. Корневая система чаще всего поверхностная, но на хорошо дренированных почвах нередко развиваются и сравнительно глубокие вертикальные ответвления (Шубин, 1973). Ствол ели обыкновенной – полнодревесный, покрыт относительно тонкой зелено-коричневой, коричневой или серой корой. Кора ели обыкновенной – гладкая, но с возрастом становится чешуйчатой и бороздчатой.
Ростовые почки небольшие – от 4 до 6 миллиметров, яйцевидно-конические, рыжие с сухими чешуйками. Репродуктивные почки более крупные и достигают 7-10 миллиметров.
Xвоя у ели обыкновенной четырехгранная, острая, темно-зеленая, жесткая, блестящая, длиной до 10-30 мм и толщиной 1-2 миллиметра. Держится на побегах 5-10 лет и опадает в течение всего года, но наиболее интенсивно с октября по май.
Ель обыкновенная цветет в мае – июне. Шишки созревают осенью на следующий год после цветения, семена выпадают в конце зимы и ранней весной следующего года. Мужские колоски удлиненной цилиндрической формы расположены на побегах прошлого года. Шишки веретенообразно цилиндрической формы, длиной от 6 до 16 и диаметром от 2,5 до 4 сантиметров, расположены на концах ветвей. Молодые шишки бывают светло-зеленые, темно фиолетовые или розоватые, а зрелые принимают иной оттенок светло-бурого или красно-бурого цвета. Зрелые шишки содержат на стержне от 100 до 200 семенных чешуек. Семенные чешуйки – одревесневшие, обратнояйцевидные, цельнокрайние, по верхнему краю мелкозазубренные, выемчатые. На каждой семенной чешуе содержатся 2 углубления для семян (Казимиров, 1983). Семена ели обыкновенной коричневого цвета, относительно небольшие, длиной от 3 до 5 миллиметров. Масса 1000 семян от 3 до 9 грамм. Всхожесть семян варьируется от 30 до 85 процентов в зависимости от условий произрастания. Условия произрастания детерминируют и наличие повторения урожайных годов, которые наступают в среднем через каждые 4-8 лет.
Ель обыкновенная – вид, произрастающий на относительно большом ареале, в различных почвенных и климатических условиях. В результате этого ель обыкновенная отличается большой внутривидовой полиморфностью (по типу ветвления, окраске шишек, строению кроны, фенологии и т.д.), следовательно, и наличием большого числа экотипов. По отношению к температуре воздуха ель обыкновенная теплолюбивая, но в тоже время – холодостойкая порода, произрастающая в зоне умеренного и прохладного климата со средней годовой температурой от -2,9 до +7,4 градусов и температурой самого теплого месяца в году от +10 до +20 градусов (Чертовской, 1978). Ареал ели обыкновенной распространяется в диапазоне от 370 до 1600 мм выпадающих осадков в год.
Вопрос увлажнения почвы тесно связан с ее аэрацией. Ель обыкновенная хоть и способна расти в условиях избыточного увлажнения, но хорошей продуктивности следует ожидать только в тех случаях, где вода проточная. На сырых почвах ель вываливается уже при скорости 6-7 метров в секунду, а на свежих и сухих почвах выдерживают потоки ветра скоростью 15 метров в секунду. Скорость ветра более 20 метров в секунду вызывает массовый повал.
Наиболее интенсивным ростом ель обыкновенная отличается на супесчаных и суглинистых почвах, подстилаемых на глубине 1-1,5 метров глинами или суглинками. Следует отметить, что строгих правил требовательности к почве ее сложению и механическому составу как таковых нет, поскольку требовательность ели к почве несет зональный характер. Ель обыкновенная имеет высокий порог толерантности к кислотности почвы и способна произрастать при колебаниях pH от 3,5 до 7,0. Ель обыкновенная относительно требовательна к минеральному питанию (Казимиров, 1983).
Учет подлеска и живого напочвенного покрова
Неоднородность качественных и количественных характеристик подроста выражается, в первую очередь, через понятие жизнеспособность подроста. Жизнеспособность подроста по «Энциклопедии лесного хозяйства» (2006) представляет собой способность молодого поколения материнского подроста существовать и функционировать в меняющихся условиях среды.
Многие исследователи, такие как И.И. Гусев (1998), М.В. Никонов (2001), В.В. Горошков (2003), В.А. Алексеев (2004), V.A. Alexeyev (1997) и другие отмечали, что изучение качественных параметров ельников, по большому счету, сводится к изучения состояния древостоев.
Состояние древостоя является последствием сложных процессов и этапов, через которые проходит растение от своего зачатка и образования семени до перехода в господствующий ярус. Этот длительный процесс метаморфозов растения требует разделения на различные этапы, каждый из которых нужно изучать в отдельном порядке.
Таким образом, можно констатировать, что понятию жизнеспособность и состояние подроста уделяется сравнительно мало внимания (Писаренко, 1977; Алексеев, 1978; Калинин, 1985; Пугачевский, 1992; Грязькин, 2000, 2001; Григорьев, 2008).
Большинство исследователей утверждает, что под пологом спелых древостоев имеется достаточное количество жизнеспособного подроста ели, однако при этом чаще всего не выявляется взаимообусловленность состояния подроста и его пространственного распределения от характеристик материнского древостоя.
Есть и исследователи не утверждающие, что под пологом материнского древостоя должен быть жизнеспособный подрост способный полноценно заменить в перспективе материнский древостой (Писаренко, 1977; Алексеев, 1978; Пугачевский, 1992).
Колебания по высоте и групповое распределение подроста ели позволили некоторым авторам утверждать, что подрост ели в целом не способен обеспечить предварительное возобновление при условии интенсивных лесозаготовительных работ (Мойланен, 2000).
Еще исследования Варгаса де Бедемара (1846) установили, что число стволов с возрастом резко снижается, и что из проросших всходов в процессе естественного отбора и дифференциации к возрасту спелости сохраняется всего лишь около 5 процентов.
Процесс дифференциации наиболее выражен в «молодости» насаждения, где в наибольшей степени выделяются угнетенные классы по состоянию, и постепенно захватывает «старость». По словам Г.Ф. Морозова, который ссылается на более ранние работы Я.С. Медведева (1910) в данном направлении, общей чертой подроста растущего в насаждении является угнетенность. Свидетельством этому является тот факт, что в возрасте 60-80 лет подрост ели под пологом очень часто не превышает 1-1,5 м, тогда как подрост ели на воле в этом же возрасте достигает высоты 10-15 метров.
Однако, Г.Ф. Морозов (1904) отмечает, что производительность и продуктивность отдельных экземпляров подроста может измениться и в лучшую сторону, стоит только изменить условия среды. Все экземпляры подроста, различной степени угнетенности, отличаются от подроста на воле по морфологическим характеристикам вегетативных органов, в т.ч. меньшим количеством почек, иной формой кроны, слабо развитой корневой системой и так далее. Такие морфологические изменения у ели, как образование зонтикообразной кроны, развивающейся в горизонтальном направлении, является приспособлением растения к максимально эффективному использованию «скудного» света, проникающего к подросту. Изучая поперечные разрезы стволиков подроста ели, растущего в условиях Ленинградского округа (Охтинская дача), Г.Ф. Морозов отмечал, что у отдельных экземпляров годичные слои были густо сомкнуты на начальном этапе жизни (что свидетельствует о степени угнетенности растения), а затем резко расширялись в результате некоторых лесохозяйственных мероприятий (в частности изреживания), изменяющих условия среды.
Подрост ели, резко оказавшись на открытом пространстве, погибает и от излишнего физиологического испарения в силу того, что на открытых участках этот процесс протекает с большей активностью, к которой подрост, растущий под пологом, не приспособлен. Чаще всего этот подрост погибает в результате резкого изменения обстановки, но, как отмечал Г. Ф. Морозов, в некоторых случаях, после долгой борьбы, он начинает оправляться и выживает. Возможность подроста выжить в таких обстоятельствах определяется рядом факторов, таких как степень его угнетенности, степень резкости изменения условий среды, и, конечно биотическими и абиотическими факторами, влияющими на рост и развитие растения.
Отдельные экземпляры подроста часто сильно различаются в пределах одного массива таким образом, что один экземпляр подроста, отмеченный до рубки как нежизнеспособный, оправился, а другой так и остался в категории нежизнеспособного. Подрост ели, образовавшийся на плодородных почвах под пологом березы или сосны, часто не реагирует на удаление верхнего яруса, т.к. световой дефицит не испытывал даже при его наличии (Cajander, 1934, Vaartaja, 1952). После буферного периода адаптации прирост подроста в высоту многократно увеличивается, но мелкий подрост требует больше времени для функциональной реструктуризации вегетативных органов (Koistinen и Valkonen, 1993).
Косвенное подтверждение факта выраженной способности подроста ели менять категорию состояния в лучшую сторону дал P. Mikola (1966), отмечая, что значительная часть забракованных ельников (исходя из состояния подроста), в процессе лесоинвентаризации Финляндии, признана позже пригодной для лесовыращивания.
Возрастная структура как индикатор состояния подроста
В зависимости от структуры насаждения под полог ельников может проникать от 3 до 17 процентов фотосинтетический активной радиации. Нужно также отметить, что по мере ухудшения эдафических условий уменьшается и степень поглощения этой радиации (Алексеев, 1975).
Средняя освещенность в нижних ярусах ельников в черничных типах леса чаще всего не превышает 10 %, а это, в свою очередь, в среднем обеспечивает минимальную энергию годового прироста, который составляет от 4 до 8 см (Чертовской, 1978).
Исследования в Ленинградской области, проведенные под руководством А.В. Грязькина (2001) показывают, что относительная освещенность на поверхности почвы под пологом древостоев составляет 0,3-2,1% от полной, а этого недостаточно для успешного роста и развития молодого поколения ели. Данные экспериментальные исследования показали, что годовой прирост молодого поколения ели увеличивается с 5 до 25 см при увеличении проникающего под полог света с 10 до 40%.
Жизнеспособный подрост ели в подавляющем большинстве случаев растет только в окнах полога елового древостоя, поскольку в окнах подрост ели не испытывает недостатка света, а к тому же напряженность корневой конкуренции там значительно ниже, нежели в приствольной части древостоя (Мелехов, 1972).
В.Н. Сукачев (1953) утверждал, что гибель подроста в большей степени определена корневой конкуренцией материнских деревьев, а только потом дефицитом света. Такое утверждение он подкреплял фактом, что на самых ранних этапах жизни подроста (первые 2 года) «происходит сильный отпад ели независимо от освещенности». Такие авторы как Е.В. Максимов (1971), В.Г. Чертовский (1978), А.В. Грязькин (2001), К.С. Бобкова (2009) и другие подвергают сомнению подобные предположения.
По данным Е.В. Максимова (1971), нежизнеспособным подрост становится при освещенности от 4 до 8 % от полной. Жизнеспособный подрост формируется в просветах между кронами взрослых деревьев, где освещенность составляет в среднем 8-20%, и характеризуется световой хвоей и хорошо развитой корневой системой. Иными словами, жизнеспособный подрост приурочен к просветам в пологе, а сильно-угнетенный подрост находится в зоне плотной сомкнутости верхних ярусов (Бобкова, 2009).
В.Г. Чертовской (1978) также утверждает, что на жизнеспособность ели решающее влияние оказывает свет. По его доводам, в среднеплотных насаждениях жизнеспособный подрост ели обычно составляет более 50-60% от общего количества. В сильно сомкнутых ельниках преобладает нежизнеспособный подрост.
Исследования в Ленинградской области показали, что режимом освещенности, т.е. сомкнутостью полога, определяется доля жизнеспособного подроста. При сомкнутости полога 0,5-0,6 преобладает подрост высотой более 1 м. При этом доля жизнеспособного подроста превышает 80%. При сомкнутости 0,9 и более (относительная освещенность менее 10%) жизнеспособный подрост чаще всего отсутствует (Грязькин, 2001).
Однако, не стоит недооценивать и другие экологические факторы, такие как структура почвы, ее влажность, а также температурный режим (Рысин, 1970; Пугачевский, 1983, Haners, 2002).
Хоть ель и относится к теневыносливым породам, подрост ели в высокоплотных насаждениях все же испытывает большие трудности при низкой освещенности. В результате качественные характеристики подроста в плотных насаждениях заметно хуже по сравнению с подростом, растущим в среднеплотных и низкоплотных насаждениях (Вялых, 1988).
По мере роста и развития подроста ели порог толерантности к низкому освещению снижается. Уже в возрасте девяти лет резко увеличивается потребность подроста ели в освещенности (Афанасьев, 1962).
От плотности древостоев зависят размер, возраст и состояние подроста. Большинство спелых и перестойных хвойных насаждений характеризуется разновозрастностью (Пугачевский, 1992). Наибольшее количество особей подроста встречается при полноте 0,6-0,7 (Атрохин, 1985, Касимов, 1967). Эти данные подтверждаются и исследованиями А.В. Грязькина (2001), которые показали, что «оптимальные условия для формирования жизнеспособного подроста численностью 3-5 тыс. экз./га складываются под пологом древостоев полнотой 0,6-0,7».
Н.Е. Декатов (1931) утверждал, что основной предпосылкой для появления жизнеспособного подроста ели в кисличном типе леса, является то, чтобы полнота материнского полога была в диапазоне 0,3-0,6.
Жизнеспособность, следовательно, и прирост в высоту во многом детерминированы плотностью насаждения, о чем свидетельствуют исследования А.В. Грязкина (2001). По данным этих исследований, прирост нежизнеспособного подроста в ельниках-кисличниках при относительной полноте древостоев 0,6 такой же, как и прирост жизнеспособного подроста, при полноте ельника кисличника 0,7-0,8.
В ельниках черничного типа леса с увеличением полноты древостоя, средняя высота подроста уменьшается и эта зависимость близка к линейной связи (Грязькин, 2001).
Исследования Н.И. Казимирова (1983) показали, что в ельниках лишайниковых полнотой 0,3-0,5 еловый подрост редкий и качественно неудовлетворительный. Совершено иначе обстоит ситуация с кисличниками, а особенно с брусничным и черничным типами леса, где несмотря на высокую сомкнутость имеется достаточное количество подроста удовлетворительного по жизненному состоянию.
Зависимость динамики состояния подроста ели от давности рубки
С увеличением относительной полноты древостоя увеличивается и доля среднего и крупного жизнеспособного подроста ели, поскольку конкуренция за свет в столь сомкнутом пологе больше всего отражается на мелком подросте. При высокой полноте древостоя доля нежизнеспособного мелкого подроста ели также весьма велика. Однако, эта доля значительно больше при небольшой относительной полноте, поскольку в таких световых условиях увеличивается конкуренция, от которой в первую очередь страдает мелкий подрост.
С увеличением относительной полноты древостоя доля мелкого нежизнеспособного подроста изменяется следующим образом: при небольшой полноте доля мелкого нежизнеспособного подроста наибольшая, затем она падает и минимума достигает при полноте 0,7, а затем снова увеличивается при увеличении полноты (рисунок 3.40).
Распределение подроста ели по категориям состояния и крупности подтверждает, что жизненный потенциал подроста выросшего в условиях Лисинского лесничества больше, чем подроста ели в Карташевском лесничестве. Это особенно четко прослеживается по высотной структуре подроста, поскольку доля среднего и крупного подроста ели, как правило, больше на лисисинских объектах при аналогичных лесорастительных условиях (рисунки 3.39-3.40).
О лучшем жизненном потенциале подроста ели на лисинских объектах свидетельствуют и темпы роста подроста, которые показаны на рисунках 3.41-42. Для каждой группы возраста вне зависимости от жизненного состояния средняя высота подроста ели на лисинских объектах больше средней высоты подроста выросшего в условиях Карташевского лесничества. Это лишний раз подтверждает тезис, что в относительно менее благоприятных условиях среды (с точки зрения влажности почвы и ее плодородия – более близких к черничному типу леса) подрост ели способен больше проявить свои конкурентные способности. Отсюда следует, что и изменения, происходящие в пологе в результате антропогенного или иного воздействия, более положительный результат дают в контексте улучшения состояния подроста ели в условиях Лисинского, нежели Карташевского лесничества.
1. На каждом этапе развития численность подроста, как и структура по высоте, по возрасту на опытных участках меняются разнонаправленно. Однако выявлена определенная закономерность: чем больше изменяется численность подроста (после урожайных семенных лет она резко увеличивается), тем сильнее меняется структура подроста по высоте и возрасту. Если при увеличении численности подроста за счет самосева происходит существенное уменьшение средней высоты и среднего возраста, то при уменьшении численности в результате отпада, средняя высота и средний возраст могут увеличиваться – если в отпад переходит преимущественно мелкий подрост, или уменьшаться – если в отпад переходит преимущественно крупный подрост.
2. За 30 лет численность подроста под пологом ельника кисличника и ельника черничника изменялась, в этом компоненте фитоценоза смена поколений идет непрерывно - основная часть старшего поколения переходит в отпад, а подрост новых генераций регулярно появляется и в первую очередь - после обильного урожая семян.
3. За три десятилетия состав подроста на объектах наблюдений изменился значительно, доля лиственных пород заметно увеличилась и достигла 31- 43 % (после проведенной рубки). В начале эксперимента она не превышала 10%.
4. На секции А экологического стационара численность подроста ели за 30 лет увеличилась на 2353 экземпляра, а с учетом сохранившихся модельных экземпляров общая численность подроста ели к 2013 году составила 2921 экз./га. В 1983 году всего было 3049 экз./га.
5. За три десятилетия под пологом ельника-черничника и ельника кисличника доля подроста, перешедшего из категории «нежизнеспособный» в категорию «жизнеспособный» составила 9 % на секции А, 11% на секции Б и 8 % на секции В, т.е. в среднем около 10%. Исходя из общей численности подроста на опытном участке 3-4 тыс./га, эта доля существенна и заслуживает внимания при проведении учетных работ при оценке успешности естественного возобновления ели в указанных типах леса. 103 6. Из категории «жизнеспособный» в категорию «нежизнеспособный» за указанный период времени перешло от 19 до 24 %, а сразу из категории «жизнеспособный» в категорию «сухой» (минуя категорию «нежизнеспособный») – от 7 до 11 %. 7. Из общего количества растущего подроста на секции А (1613 экземпляров) перешло в отпад 1150 экземпляров подроста разной высоты и разного возраста, т.е. около 72 %. На секции Б – 60 %, а на секции В – 61 %. 8. В ходе наблюдений доля сухого подроста увеличивалась с увеличением высоты и возраста модельных экземпляров. Если в 1983-1989 гг. она составляла 6,3-8,0 % от общего количества, то к 2013 году к сухому подросту относилось уже от 15 (ельник черничник) до 18-19 % (ельник кисличник). 9. Из общего количества паспортизированного подроста на секции А, деревьями перечетных размеров стали 127 экземпляра, т.е. 7,3 %. Из них большую часть (4,1 %) составляют те экземпляры, которые перешли в разные годы из категории «нежизнеспособный» в категорию «жизнеспособный». 10. Многократность учета одних и тех же экземпляров подроста ели в течение длительного периода времени позволяет указать на основные причины переходов из категории «нежизнеспособный» в категорию «жизнеспособный». 11. Изменение структуры подроста по высоте и по возрасту, колебание численности – процесс динамичный, при котором одновременно совмещается два взаимно противоположных процесса: отпад и приход новых поколений подроста. 12. Переходы подроста из одной категории состояния в другую, как правило, чаще происходит среди мелкого подроста. Чем меньше возраст подроста, тем вероятнее позитивный переход. Если за первые 6 лет наблюдений из категории «НЖ» перешло в категорию «Ж» около 3 % экз. (при среднем возрасте подроста 19 лет), то через 20 лет – менее 1%, а через 30 лет – всего 0,2%. 13. Динамика состояния подроста выражена и по типам леса. Переходы нежизнеспособного подроста в категорию «жизнеспособный» более вероятны в ельнике черничнике, чем в ельнике кисличнике.

