ب. ساختار اسید دئوکسی ریبونوکلئیک (DNA). اسیدهای نوکلئیک: تاریخچه تحقیقات، توصیف خواص شیمیایی DNA
وزارت آموزش و پرورش و علوم فدراسیون روسیه
موسسه آموزشی خودمختار ایالتی فدرال
آموزش عالی
"دانشگاه ملی پژوهشی فناوری کازان"
انستیتوی مهندسی صنایع غذایی
بخش بیوتکنولوژی مواد غذایی
چکیده در مورد موضوع
اسیدهای نوکلئیک. DNA و RNA
تکمیل شده توسط: Radenko V.
گروه 625 M-52
اسیدهای نوکلئیک -ترکیبات آلی طبیعی با مولکولی بالا که ذخیره و انتقال اطلاعات ارثی (ژنتیکی) را در موجودات زنده تضمین می کند. هر موجود زنده دارای 2 نوع اسید نوکلئیک است: اسید ریبونوکلئیک (RNA) و اسید دئوکسی ریبونوکلئیک (DNA). وزن مولکولی کوچکترین اسید نوکلئیک شناخته شده، RNA انتقالی (tRNA)، تقریباً 25 کیلو دالتون است. DNA بزرگترین مولکول پلیمری است. وزن مولکولی آنها از 1000 تا 1000000 کیلو دالتون متغیر است. DNA و RNA از واحدهای مونومر - نوکلئوتیدها تشکیل شده است، به همین دلیل است که اسیدهای نوکلئیک را پلی نوکلئوتید می نامند.
ساختار نوکلئوتیدها
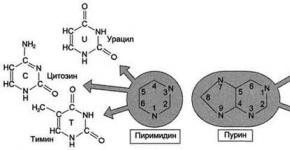
هر نوکلئوتید شامل 3 جزء شیمیایی متفاوت است: یک باز نیتروژن دار هتروسیکلیک، یک مونوساکارید (پنتوز) و یک باقیمانده اسید فسفریک. بسته به تعداد باقی مانده های اسید فسفریک موجود در مولکول، مونوفسفات های نوکلئوزیدی (NMP)، نوکلئوزید دی فسفات ها (NDP) و نوکلئوزید تری فسفات ها (NTP) متمایز می شوند (شکل 4-1). اسیدهای نوکلئیک حاوی دو نوع باز نیتروژن هستند: پورین - آدنین(آ)، گوانین(G) و پیریمیدین - سیتوزین(با)، تیمین(T) و اوراسیل(U). شماره گذاری اتم ها در پایه ها در داخل چرخه نوشته شده است (شکل 4-2). پنتوزها در نوکلئوتیدها یا ریبوز (در RNA) یا دئوکسی ریبوز (در DNA) هستند. برای تشخیص تعداد اتم ها در پنتوزها از شماره گذاری اتم ها در پایه، ضبط در قسمت بیرونی چرخه انجام می شود و یک عدد اول (") به عدد اضافه می شود - 1، 2، 3، 4 و 5" (شکل 4-3). پنتوز به پایه متصل می شود پیوند N-گلیکوزیدی،توسط اتم C 1 پنتوز (ریبوز یا دئوکسی ریبوز) و اتم N 1 پیریمیدین یا اتم N 9 پورین تشکیل شده است (شکل 4-4). نوکلئوتیدهایی که در آنها پنتوز با ریبوز نشان داده می شود، ریبونوکلئوتید و اسیدهای نوکلئیک ساخته شده از ریبونوکلئوتیدها، اسیدهای ریبونوکلئیک یا RNA نامیده می شوند. اسیدهای نوکلئیک که مونومرهای آنها شامل دئوکسی ریبوز است، اسیدهای دئوکسی ریبونوکلئیک یا DNA نامیده می شوند. اسیدهای نوکلئیک با توجه به ساختار آنها به عنوان دسته بندی می شوند
برنج. 4-1. نوکلئوزیدهای مونو، دی و تری فسفات آدنوزین.نوکلئوتیدها استرهای فسفر نوکلئوزیدها هستند. باقی مانده اسید فسفریک به اتم کربن 5 اینچی پنتوز (پیوند فسفواستر 5 اینچی) متصل می شود.

برنج. 4-2. پایه های پورین و پیریمیدین.

برنج. 4-3. پنتوزها 2 نوع وجود دارد - β-D-ریبوز در ترکیب نوکلئوتیدهای RNA و β-D-2-دئوکسی ریبوز در ترکیب نوکلئوتیدهای DNA.
کلاس پلیمرهای خطی ستون فقرات اسید نوکلئیک دارای ساختار یکسانی در تمام طول مولکول است و از گروه های متناوب - پنتوز-فسفات-پنتوز- تشکیل شده است (شکل 4-5). گروه های متغیر در زنجیره های پلی نوکلئوتیدی، بازهای نیتروژنی هستند - پورین ها و پیریمیدین ها. مولکول های RNA شامل آدنین (A)، اوراسیل (U)، گوانین (G) و سیتوزین (C) هستند، در حالی که DNA حاوی آدنین (A)، تیمین (T)، گوانین (G) و سیتوزین (C) است. ساختار منحصر به فرد و فردیت عملکردی مولکول های DNA و RNA توسط ساختار اولیه آنها تعیین می شود - دنباله ای از بازهای نیتروژنی در زنجیره پلی نوکلئوتیدی.

برنج. 4-4. نوکلئوتیدهای پورین و پیریمیدین.

برنج. 4-5. قطعه ای از زنجیره DNA
ب. ساختار اسید دئوکسی ریبونوکلئیک (DNA)
ساختار اولیه DNA -ترتیب تناوب مونوفسفات های دئوکسی ریبونوکلئوزیدی (dNMP) در زنجیره پلی نوکلئوتیدی. هر گروه فسفات در زنجیره پلی نوکلئوتیدی، به استثنای باقیمانده فسفر در انتهای 5 اینچی مولکول، در تشکیل دو پیوند استری شامل اتمهای کربن 3 اینچی و 5 اینچی دو دئوکسی ریبوز همسایه شرکت میکند، بنابراین پیوند بین مونومرها با فسفودی استر 3" 5" مشخص می شوند. نوکلئوتیدهای انتهایی DNA از نظر ساختار متمایز می شوند: در انتهای 5 اینچ یک گروه فسفات و در انتهای 3 اینچ زنجیره یک گروه آزاد OH وجود دارد. به انتهای آن ها انتهای 5 و 3 می گویند. دنباله خطی دئوکسی ریبونوکلئوتیدها در زنجیره پلیمر DNA معمولاً با استفاده از یک کد تک حرفی مخفف می شوند، به عنوان مثال -A-G-C-T-T-A-C-A- از انتهای 5 اینچ تا 3 اینچ.
هر مونومر اسید نوکلئیک حاوی یک باقیمانده اسید فسفریک است. در PH 7 گروه فسفات به طور کامل یونیزه می شود، بنابراین in vivoاسیدهای نوکلئیک به صورت پلی آنیون (دارای بارهای منفی متعدد) وجود دارند. بقایای پنتوز نیز خاصیت آبدوستی از خود نشان می دهند. بازهای نیتروژن تقریباً در آب نامحلول هستند، اما برخی از اتم های حلقه های پورین و پیریمیدین قادر به تشکیل هستند. پیوند های هیدروژنی.
ساختار ثانویه DNAدر سال 1953، J. Watson و F. Crick مدلی از ساختار فضایی DNA ارائه کردند. بر اساس این مدل، مولکول DNA شکل یک مارپیچ دارد که توسط دو زنجیره پلی نوکلئوتیدی که نسبت به یکدیگر و حول یک محور مشترک پیچ خورده اند، تشکیل شده است. مارپیچ دوتایی راست دست،زنجیره پلی نوکلئوتیدی در آن است ضد موازی(شکل 4-6)، یعنی. اگر یکی از آنها در جهت 3"→5" باشد، دومی در جهت 5"→3" است. بنابراین، در هر پایان
برنج. 4-6. مارپیچ دوگانه DNA.
مولکول های DNA از دو رشته ضد موازی با یک توالی نوکلئوتیدی مکمل تشکیل شده اند. زنجیرها نسبت به یکدیگر به شکل یک مارپیچ راست دست پیچ خورده اند به طوری که تقریباً 10 جفت نوکلئوتید در هر چرخش مولکول وجود دارد. تمام پایه های رشته های DNA در داخل مارپیچ دوگانه قرار دارند و ستون فقرات پنتوز فسفات در خارج است. زنجیره های پلی نوکلئوتیدی به دلیل پیوندهای هیدروژنی بین بازهای نیتروژن دار مکمل پورین و پیریمیدین A و T (دو پیوند) و بین G و C (سه پیوند) نسبت به یکدیگر نگه داشته می شوند (شکل 4-7). با این ترکیب، هر جفت شامل سه حلقه است، بنابراین اندازه کلی این جفتهای پایه در تمام طول مولکول یکسان است.

برنج. 4-7. جفت بازهای پورین- پیریمیدین در DNA.
پیوندهای هیدروژنی با سایر ترکیبات پایه در یک جفت ممکن است، اما آنها بسیار ضعیف تر هستند. توالی نوکلئوتیدی یک زنجیره کاملا مکمل توالی نوکلئوتیدی زنجیره دوم است. بنابراین، طبق قانون چارگاف (اروین چارگاف در سال 1951 الگوهایی را در نسبت بازهای پورین و پیریمیدین در یک مولکول DNA ایجاد کرد)، تعداد بازهای پورینی (A + G) برابر با تعداد بازهای پیریمیدینی (T + C) است. . پایه های مکمل در هسته مارپیچ چیده شده اند. بین پایه های یک مولکول دو رشته ای در یک پشته، فعل و انفعالات آبگریز،تثبیت مارپیچ دوگانه
این ساختار از تماس باقی مانده های نیتروژنی با آب جلوگیری می کند، اما پشته پایه ها نمی تواند کاملاً عمودی باشد. جفت های پایه کمی از یکدیگر فاصله دارند. در ساختار تشکیل شده، دو شیار متمایز می شوند - یک شیار بزرگ به عرض 2.2 نانومتر و یک شیار کوچک به عرض 1.2 نانومتر. پایه های نیتروژنی در ناحیه شیارهای اصلی و فرعی با پروتئین های خاصی که در سازماندهی ساختار کروماتین دخیل هستند تعامل دارند.
ساختار سوم DNA (ابر مارپیچ DNA)هر مولکول DNA در یک کروموزوم جداگانه بسته بندی می شود. سلول های دیپلوئید انسانی حاوی 46 کروموزومطول کل DNA همه کروموزوم های یک سلول 1.74 متر است، اما در هسته ای بسته بندی شده است که قطر آن میلیون ها بار کوچکتر است. برای قرار دادن DNA در هسته سلول، ساختار بسیار فشرده ای باید تشکیل شود. فشردگی و ابرپیچشدن DNA با استفاده از انواع پروتئینهایی که با توالیهای خاصی در ساختار DNA تعامل دارند، انجام میشود. تمام پروتئین هایی که به DNA یوکاریوتی متصل می شوند را می توان به 2 گروه تقسیم کرد: گیسگون و پروتئین های غیر هیستونی.مجموعه پروتئین ها با DNA هسته سلول ها کروماتین نامیده می شود.
هیستون ها- پروتئین هایی با وزن مولکولی 11-21 کیلو دالتون حاوی مقادیر زیادی آرژنین و لیزین. هیستون ها به دلیل بار مثبت خود، پیوندهای یونی با گروه های فسفات با بار منفی که در قسمت بیرونی مارپیچ دوگانه DNA قرار دارند، تشکیل می دهند. 5 نوع هیستون وجود دارد. چهار هیستون H2A، H2B، H3 و H4 یک کمپلکس پروتئین اکتامری (H2A، H2B، H3، H4) را تشکیل می دهند که به نام "هسته نوکلئوزومی"(از انگلیسی هسته نوکلئوزومی). مولکول DNA روی سطح اکتامر هیستون «باد» میزند و 1.75 چرخش (حدود 146 جفت نوکلئوتید) را تکمیل میکند. این مجموعه از پروتئین های هیستون با DNA به عنوان واحد ساختاری اصلی کروماتین عمل می کند. "نوکلئوزوم". DNA اتصال دهنده ذرات نوکلئوزومی DNA پیوند دهنده نامیده می شود. به طور متوسط، DNA پیوند دهنده 60 جفت باقیمانده نوکلئوتیدی است. مولکول های هیستون H1 به DNA در نواحی بین هسته ای (توالی های پیوند دهنده) متصل می شوند و از این نواحی در برابر عمل نوکلئازها محافظت می کنند (شکل 4-8).
 برنج. 4-8. ساختار نوکلئوزومی
برنج. 4-8. ساختار نوکلئوزومی
هشت مولکول هیستون (H2A، H2B، H3، H4) 2 هسته نوکلئوزوم را تشکیل می دهند که DNA تقریباً 1.75 دور آن را تشکیل می دهد. DNA باقیماندههای اسید آمینه لیزین، آرژنین و گروههای آمینه انتهایی هیستونها را میتوان تغییر داد: استیله، فسفریله، متیله، یا با پروتئین یوبیکوئیتین (پروتئین غیر هیستونی) در تعامل است. تغییرات میتوانند برگشتپذیر یا غیرقابل برگشت باشند؛ آنها بار و ترکیب هیستونها را تغییر میدهند و این بر تعامل هیستونها با یکدیگر و با DNA تأثیر میگذارد. فعالیت آنزیم های مسئول تغییرات تنظیم شده و به مرحله چرخه سلولی بستگی دارد. تغییرات، بازآرایی ساختاری کروماتین را ممکن میسازد.
پروتئین های کروماتین غیر هیستونیهسته یک سلول یوکاریوتی حاوی صدها پروتئین غیرهیستونی مختلف متصل به DNA است. هر پروتئین مکمل توالی خاصی از نوکلئوتیدهای DNA است (سایت DNA).این گروه شامل خانواده پروتئینهای مخصوص سایت از نوع «انگشت روی» است (به بخش 1 مراجعه کنید). هر "انگشت روی" یک محل خاص متشکل از 5 جفت نوکلئوتید را تشخیص می دهد. خانواده دیگری از پروتئین های مخصوص سایت، همودایمرها هستند. قطعه ای از چنین پروتئینی در تماس با DNA دارای ساختار مارپیچ-گردش-مارپیچ است (به بخش 1 مراجعه کنید). گروهی از پروتئین های ساختاری و تنظیمی که به طور مداوم با کروماتین در ارتباط هستند شامل پروتئین های با تحرک بالا می باشد. پروتئین های HMG- از انگلیسی، پروتئین های ژل با تحرک بالا). آنها وزن مولکولی کمتر از 30 کیلو دالتون دارند و با محتوای بالای آمینو اسیدهای باردار مشخص می شوند. پروتئین های HMG به دلیل وزن مولکولی کم، تحرک بالایی در طول الکتروفورز ژل پلی آکریل آمید دارند. پروتئین های غیر هیستونی نیز شامل آنزیم های همانندسازی، رونویسی و ترمیم می شوند. با مشارکت پروتئینهای ساختاری، تنظیمکننده و آنزیمهای دخیل در سنتز DNA و RNA، رشته نوکلئوزومها به مجموعهای بسیار متراکم از پروتئینها و DNA تبدیل میشوند. ساختار حاصل 10000 برابر کوتاهتر از مولکول DNA اصلی است.
سه ماکرومولکول اصلی در یک موجود زنده وجود دارد: پروتئین ها و دو نوع اسید نوکلئیک. به لطف آنها، فعالیت حیاتی و عملکرد صحیح کل بدن حفظ می شود. اسیدهای نوکلئیک چیست؟ چرا آنها مورد نیاز هستند؟ بیشتر در این مورد در ادامه مقاله.
اطلاعات کلی
اسید نوکلئیک یک بیوپلیمر، یک ترکیب آلی با مولکولی بالا است که توسط بقایای نوکلئوتیدی تشکیل می شود. انتقال تمام اطلاعات ژنتیکی از نسلی به نسل دیگر وظیفه اصلی اسیدهای نوکلئیک است. ارائه زیر این مفهوم را با جزئیات بیشتری توضیح خواهد داد.
تاریخچه مطالعه
اولین نوکلئوتید مورد مطالعه در سال 1847 از ماهیچه گاو جدا شد و "اسید اینوزینیک" نام گرفت. در نتیجه مطالعه ساختار شیمیایی، مشخص شد که این یک ریبوزید-5'-فسفات و حاوی پیوند N-گلیکوزیدی است.در سال 1868، ماده ای به نام "نوکلئین" کشف شد. این ماده توسط شیمیدان سوئیسی، فردریش میشر، در حین تحقیق بر روی برخی از مواد بیولوژیکی کشف شد. این ماده شامل فسفر بود. این ترکیب دارای خواص اسیدی بود و تحت تأثیر آنزیم های پروتئولیتیک تجزیه نمی شد.

این ماده فرمول C29H49N9O22P3 را دریافت کرد.فرض در مورد مشارکت نوکلئین در فرآیند انتقال اطلاعات ارثی در نتیجه کشف شباهت ترکیب شیمیایی آن با کروماتین مطرح شد. این عنصر جزء اصلی کروموزوم ها می باشد اصطلاح نوکلئیک اسید اولین بار در سال 1889 توسط ریچارد آلتمن معرفی شد. او بود که نویسنده روش تولید این مواد بدون ناخالصی های پروتئینی شد.لوین و جاکوب در طول مطالعه هیدرولیز قلیایی اسیدهای نوکلئیک، اجزای اصلی محصولات این فرآیند را شناسایی کردند. معلوم شد که آنها نوکلئوتیدها و نوکلئوزیدها هستند. در سال 1921، لوین پیشنهاد کرد که DNA دارای ساختار تترانوکلئوتیدی است. اما این فرضیه تایید نشد و معلوم شد که اشتباه است.

طبقه بندی
اسیدهای نوکلئیک در دو نوع وجود دارند: DNA و RNA. حضور آنها در سلول های همه موجودات زنده یافت می شود. DNA عمدتاً در هسته سلول یافت می شود. RNA در سیتوپلاسم یافت می شود. در سال 1935، طی قطعه قطعه شدن نرم DNA، 4 نوکلئوتید تشکیل دهنده DNA به دست آمد. این اجزا در حالت کریستالی ارائه می شوند. در سال 1953، واتستون و کریک تشخیص دادند که DNA دارای یک مارپیچ دوگانه است.
روش های انتخاب

روش های مختلفی برای به دست آوردن ترکیبات از منابع طبیعی ایجاد شده است. شرایط اصلی این روش ها جداسازی موثر اسیدهای نوکلئیک و پروتئین ها، کمترین تکه تکه شدن مواد به دست آمده در طول فرآیند است. امروزه روش کلاسیک بسیار مورد استفاده قرار می گیرد. ماهیت این روش تخریب دیواره های مواد بیولوژیکی و تصفیه بیشتر آنها با مواد شوینده آنیونی است. نتیجه یک رسوب پروتئین است، در حالی که اسیدهای نوکلئیک در محلول باقی می مانند. روش دیگری نیز استفاده می شود. در این حالت، اسیدهای نوکلئیک را می توان با استفاده از اتانول و سالین به حالت ژل رسوب داد. هنگام انجام این کار باید کمی احتیاط کرد. به ویژه، اتانول باید با احتیاط زیاد به محلول نمک اضافه شود تا رسوب ژل به دست آید. اسید نوکلئیک در چه غلظتی آزاد می شود، چه ناخالصی هایی در آن وجود دارد، با روش اسپکتروفتومتری قابل تشخیص است. اسیدهای نوکلئیک به راحتی توسط نوکلئازها که دسته خاصی از آنزیم ها هستند تجزیه می شوند. با چنین جداسازی، لازم است که تجهیزات آزمایشگاهی تحت درمان اجباری با مهارکننده ها قرار گیرند. اینها شامل، برای مثال، یک مهارکننده DEPC است که در جداسازی RNA استفاده می شود.
مشخصات فیزیکی
اسیدهای نوکلئیک حلالیت خوبی در آب دارند، اما تقریباً در ترکیبات آلی نامحلول هستند. علاوه بر این، آنها به خصوص به دما و سطوح pH حساس هستند. مولکول های اسید نوکلئیک با وزن مولکولی بالا می توانند توسط نوکلئاز تحت تأثیر نیروهای مکانیکی تکه تکه شوند. اینها شامل مخلوط کردن محلول و تکان دادن آن است.
اسیدهای نوکلئیک. ساختار و توابع

اشکال پلیمری و مونومری ترکیبات مورد بحث در سلول ها یافت می شود. به اشکال پلیمری پلی نوکلئوتید می گویند. در این شکل، زنجیره های نوکلئوتیدی توسط یک باقیمانده اسید فسفریک به هم متصل می شوند. با توجه به محتوای دو نوع مولکول هتروسیکلیک به نام های ریبوز و دئوکسی ریبوز، اسیدها به ترتیب اسیدهای ریبونوکلئیک و اسیدهای دی اکسی ریبونوکلئیک هستند. با کمک آنها، ذخیره، انتقال و اجرای اطلاعات ارثی رخ می دهد. از اشکال مونومر اسیدهای نوکلئیک، محبوب ترین اسید آدنوزین تری فسفریک است. در سیگنال دهی و تامین ذخایر انرژی در سلول نقش دارد.
DNA
دئوکسی ریبونوکلئیک اسید یک ماکرومولکول است. با کمک آن، فرآیند انتقال و پیاده سازی اطلاعات ژنتیکی اتفاق می افتد. این اطلاعات برای رشد و عملکرد یک موجود زنده ضروری است. در حیوانات، گیاهان و قارچ ها، DNA بخشی از کروموزوم های واقع در هسته سلول است و همچنین در میتوکندری ها و پلاستیدها یافت می شود. در باکتری ها و باستانی ها، مولکول اسید دئوکسی ریبونوکلئیک از داخل به غشای سلولی می چسبد. در چنین موجوداتی، عمدتاً مولکول های DNA حلقوی وجود دارند. آنها "پلاسمید" نامیده می شوند. دئوکسی ریبونوکلئیک اسید با توجه به ساختار شیمیایی خود یک مولکول پلیمری متشکل از نوکلئوتیدها است. این اجزا به نوبه خود حاوی یک پایه نیتروژن دار، قند و یک گروه فسفات هستند. به دلیل دو عنصر آخر است که بین نوکلئوتیدها پیوند ایجاد می شود و زنجیره ایجاد می کند. اساساً ماکرومولکول DNA به شکل یک مارپیچ از دو زنجیره ارائه می شود.

RNA
ریبونوکلئیک اسید یک زنجیره طولانی است که از نوکلئوتیدها تشکیل شده است. آنها حاوی یک پایه نیتروژن دار، قند ریبوز و یک گروه فسفات هستند. اطلاعات ژنتیکی با استفاده از دنباله ای از نوکلئوتیدها رمزگذاری می شود. RNA برای برنامه ریزی سنتز پروتئین استفاده می شود. اسید ریبونوکلئیک در طول رونویسی ایجاد می شود. این فرآیند سنتز RNA روی یک الگوی DNA است. این با مشارکت آنزیم های خاص رخ می دهد. آنها RNA پلیمراز نامیده می شوند. پس از این، اسیدهای ریبونوکلئیک الگو در فرآیند ترجمه شرکت می کنند. به این ترتیب سنتز پروتئین در ماتریس RNA اتفاق می افتد. ریبوزوم ها در این فرآیند نقش فعالی دارند. RNA های باقیمانده برای تکمیل رونویسی تحت دگرگونی های شیمیایی قرار می گیرند. در نتیجه تغییراتی که رخ می دهد، ساختارهای ثانویه و سوم اسید ریبونوکلئیک تشکیل می شود. آنها بسته به نوع RNA عمل می کنند.
محتوای مقاله
اسیدهای نوکلئیک- مولکولهای پلیمری بیولوژیکی که تمام اطلاعات مربوط به یک موجود زنده را ذخیره میکند و رشد و نمو آن را تعیین میکند و همچنین ویژگیهای ارثی را که به نسل بعدی منتقل میشود. اسیدهای نوکلئیک در هسته سلولی همه موجودات گیاهی و جانوری یافت می شوند که نام آنها را تعیین کرده است (lat. . هسته - هسته).
ترکیب زنجیره پلیمری اسیدهای نوکلئیک.
زنجیره پلیمری اسیدهای نوکلئیک از قطعات اسید فسفریک H 3 PO 3 و قطعاتی از مولکول های هتروسیکلیک که مشتقات فوران هستند، مونتاژ می شود. فقط دو نوع اسید نوکلئیک وجود دارد که هر کدام بر اساس یکی از دو نوع هتروسیکل - ریبوز یا دئوکسی ریبوز ساخته شده اند (شکل 1).
برنج. 1. ساختار ریبوز و دی اکسی ریبوز.
نام ریبوز (از لات. . دنده - دنده، گیره کاغذ) دارای پایان - ose است، که نشان می دهد که به کلاس قندها (به عنوان مثال، گلوکز، فروکتوز) تعلق دارد. ترکیب دوم گروه OH (گروه هیدروکسی) ندارد که با رنگ قرمز در ریبوز مشخص شده است. از این نظر، ترکیب سه گانه را دئوکسی ریبوز می نامند، یعنی ریبوز فاقد گروه اکسی.
زنجیره پلیمری که از قطعات ریبوز و اسید فسفریک ساخته شده است، اساس یکی از اسیدهای نوکلئیک - اسید ریبونوکلئیک (RNA) است. اصطلاح "اسید" در نام این ترکیب به این دلیل استفاده می شود که یکی از گروه های اسیدی OH اسید فسفریک بدون جانشین باقی می ماند که به کل ترکیب یک ویژگی کمی اسیدی می دهد. اگر به جای ریبوز، دئوکسی ریبوز در تشکیل زنجیره پلیمری نقش داشته باشد، اسید دئوکسی ریبونوکلئیک تشکیل می شود که نام اختصاری معروف DNA معمولاً پذیرفته شده است.
ساختار DNA
مولکول DNA به عنوان نقطه شروع در روند رشد و تکامل ارگانیسم عمل می کند. در شکل شکل 2 نشان می دهد که چگونه دو نوع ترکیب شروع متناوب در یک زنجیره پلیمری ترکیب می شوند؛ نه روش سنتز، بلکه نمودار اصلی مونتاژ یک مولکول DNA را نشان می دهد.

در نسخه نهایی، مولکول DNA پلیمری حاوی هتروسیکل های حاوی نیتروژن در قاب کناری است. چهار نوع از این ترکیبات در تشکیل DNA نقش دارند، دو نوع از آنها چرخه های شش عضوی و دو نوع چرخه متراکم هستند که در آن یک حلقه شش عضوی به یک حلقه پنج عضوی ذوب می شود (شکل 3).

برنج. 3. ساختار هتروسیکل های حاوی نیتروژنکه بخشی از DNA هستند
در مرحله دوم مونتاژ، ترکیبات هتروسیکلیک حاوی نیتروژن نشان داده شده در بالا به گروه های OH آزاد دئوکسی ریبوز اضافه می شوند و آویزهای جانبی را روی زنجیره پلیمری تشکیل می دهند (شکل 4).

مولکول های آدنین، تیمین، گوانین و سیتوزین متصل به زنجیره پلیمری با حروف اول نام ترکیبات اصلی مشخص می شوند، یعنی: آ, تی, جیو سی.
زنجیره پلیمری DNA خود دارای جهت خاصی است - هنگام حرکت ذهنی در امتداد مولکول در جهت جلو و معکوس، گروه های مشابهی که زنجیره را تشکیل می دهند در طول مسیر در توالی های مختلف مواجه می شوند. هنگام حرکت در یک جهت از یک اتم فسفر به اتم دیگر، ابتدا در طول مسیر یک گروه CH 2 و سپس دو گروه CH وجود دارد (اتم های اکسیژن را می توان نادیده گرفت)؛ هنگامی که در جهت مخالف حرکت می کنید، دنباله این گروه ها خواهد بود. معکوس شد (شکل 5).

برنج. 5. جهت گیری زنجیره پلیمری DNA. هنگام توصیف ترتیبی که هتروسیکل های متصل شده متناوب می شوند، مرسوم است که از جهت مستقیم استفاده شود، یعنی از گروه CH 2 به گروه های CH.
مفهوم "جهت رشته" به درک نحوه چیدمان دو رشته DNA در هنگام ترکیب شدن کمک می کند و همچنین مستقیماً با سنتز پروتئین مرتبط است.
در مرحله بعد، دو مولکول DNA با هم ترکیب می شوند، به طوری که ابتدا و انتهای زنجیره ها در جهت مخالف قرار می گیرند. در این حالت، هتروسیکل های دو زنجیره روبروی یکدیگر قرار دارند و به روشی بهینه قرار می گیرند، به این معنی که پیوندهای هیدروژنی بین جفت گروه های C=O و NH 2 و همچنین بین є N و NH= ایجاد می شوند که بخشی هستند. از هتروسیکل ها ( سانتی متر. پیوند هیدروژنی). در شکل شکل 6 نشان می دهد که چگونه دو زنجیره نسبت به یکدیگر قرار گرفته اند و چگونه پیوندهای هیدروژنی بین هتروسیکل ها ایجاد می شود. مهمترین جزئیات این است که جفت های متصل شده توسط پیوندهای هیدروژنی کاملاً مشخص هستند: قطعه آهمیشه با تی، و قطعه جی- همیشه با سی. هندسه کاملاً تعریف شده این گروه ها منجر به این واقعیت می شود که این جفت ها به طور بسیار دقیق با یکدیگر (مانند کلید یک قفل)، یک جفت مطابقت دارند. A-Tبا دو پیوند هیدروژنی و جفت به هم متصل می شوند G-C- سه اتصال

پیوندهای هیدروژنی به طور قابل توجهی ضعیف تر از پیوندهای ظرفیت معمولی هستند، اما به دلیل تعداد زیاد آنها در کل مولکول پلیمر، اتصال دو زنجیره کاملاً قوی می شود. یک مولکول DNA شامل ده ها هزار گروه است آ, تی, جیو سیو ترتیب تناوب آنها در یک مولکول پلیمری می تواند متفاوت باشد، برای مثال، در یک بخش معینی از زنجیره، دنباله می تواند به صورت زیر باشد: آ-آ-تی-جی-سی-جی-آ-تی-. از آنجایی که گروه های برهم کنش کاملاً تعریف شده اند، بخش مخالف مولکول پلیمر دوم لزوماً دنباله ای خواهد داشت - تی-تی-آ-سی-جی-سی-تی-آ-. بنابراین با دانستن ترتیب چینش هتروسیکل ها در یک زنجیره می توان قرار گرفتن آنها را در زنجیره دیگر نشان داد. از این متناظر نتیجه می شود که تعداد کل گروه ها در یک مولکول DNA مضاعف آبرابر تعداد گروه ها تیو تعداد گروه ها جی- تعداد سی(قاعده ی چارگاف).
دو مولکول DNA که توسط پیوندهای هیدروژنی متصل شده اند در شکل نشان داده شده است. 5 به شکل دو زنجیر تخت خوابیده، اما در واقع آنها به طور متفاوتی چیده شده اند. جهت واقعی در فضا همه پیوندها، که توسط زوایای پیوند و فعل و انفعالات هیدروژنی منقبض تعیین می شود، منجر به خمش خاصی از زنجیره های پلیمری و چرخش صفحه هتروسیکل می شود که تقریباً در اولین قطعه ویدیویی شکل نشان داده شده است. 7 با استفاده از فرمول ساختاری. کل ساختار فضایی را می توان با دقت بسیار بیشتری تنها با کمک مدل های سه بعدی منتقل کرد (شکل 7، قطعه ویدیوی دوم). در این مورد، یک تصویر پیچیده ایجاد می شود، بنابراین مرسوم است که از تصاویر ساده شده استفاده شود، که به ویژه هنگام به تصویر کشیدن ساختار اسیدهای نوکلئیک یا اسیدهای نوکلئیک به طور گسترده استفاده می شود. پروتئین ها. در مورد اسیدهای نوکلئیک، زنجیره های پلیمری به شکل نوارهای مسطح و گروه های هتروسیکلیک نشان داده می شوند. آ, تی, جیو سی- به شکل میله های جانبی یا ضربات ظرفیتی ساده، دارای رنگ های مختلف، یا در انتها حاوی حروف هتروسیکل های مربوطه (شکل 7، بخش سوم ویدیو).

هنگامی که کل ساختار حول محور عمودی می چرخد (شکل 8)، شکل مارپیچ دو مولکول پلیمر به وضوح قابل مشاهده است، گویی روی سطح استوانه زخمی شده است؛ این مارپیچ دوگانه شناخته شده DNA است.

با چنین تصویر ساده شده، اطلاعات اصلی ناپدید نمی شوند - ترتیب تناوب گروه بندی آ, تی, جیو سی، که فردیت هر موجود زنده را تعیین می کند، تمام اطلاعات در یک کد چهار حرفی ثبت می شود.
ساختار زنجیره پلیمری و حضور اجباری چهار نوع هتروسیکل برای همه نمایندگان دنیای زنده یکسان است. همه حیوانات و گیاهان عالی دارای تعداد جفت هستند آ – تیهمیشه تا حدودی بیشتر از یک زوج جی – سی. تفاوت بین DNA پستانداران و DNA گیاهان این است که پستانداران دارای یک جفت هستند آ – تیدر طول کل زنجیره کمی بیشتر (تقریباً 1.2 برابر) از جفت رخ می دهد. جی – سی. در مورد گیاهان، اولویت برای جفت اول بسیار بیشتر است (تقریباً 1.6 برابر).
DNA یکی از بزرگترین مولکول های پلیمری است که امروزه شناخته شده است؛ در برخی موجودات، زنجیره پلیمری آن از صدها میلیون واحد تشکیل شده است. طول چنین مولکولی به چندین سانتی متر می رسد که برای اجسام مولکولی مقدار بسیار زیادی است. زیرا از آنجایی که سطح مقطع مولکول تنها 2 نانومتر است (1 نانومتر = 10-9 متر)، نسبت های آن را می توان با ریل راه آهن به طول ده ها کیلومتر مقایسه کرد.
خواص شیمیایی DNA
در آب، DNA محلولهای چسبناکی را تشکیل میدهد؛ وقتی چنین محلولهایی تا دمای 60 درجه سانتیگراد گرم میشوند یا در معرض مواد قلیایی قرار میگیرند، مارپیچ دوگانه به دو زنجیره تبدیل میشود که اگر به شرایط اولیه برگردیم، دوباره میتوانند با هم متحد شوند. در شرایط کمی اسیدی، هیدرولیز اتفاق میافتد، در نتیجه قطعات –P-O-CH2 تا حدی شکسته میشوند و به ترتیب قطعات –P-OH و HO-CH2 را تشکیل میدهند که منجر به تشکیل مونومر، دایمر (دوبل) میشود. ) یا اسیدهای تریمریک (سه گانه)، که پیوندهایی هستند که زنجیره DNA از آنها جمع شده است (شکل 9).
برنج. 9. قطعات به دست آمده با برش DNA.
هیدرولیز عمیق تر، جداسازی بخش های دئوکسی ریبوز از اسید فسفریک و همچنین گروه را ممکن می سازد. جیاز دئوکسی ریبوز، به عنوان مثال، مولکول DNA را با جزئیات بیشتر به اجزای تشکیل دهنده آن جدا می کند. تحت تأثیر اسیدهای قوی (علاوه بر تجزیه قطعات -P(O)-O-CH 2 -)، گروه ها نیز از هم جدا می شوند. آو جی. عمل سایر معرف ها (مثلا هیدرازین) امکان جداسازی گروه ها را فراهم می کند تیو سی. برش ظریف تر DNA به اجزاء با استفاده از یک آماده سازی بیولوژیکی - دئوکسی ریبونوکلئاز، جدا شده از پانکراس انجام می شود (پایان - آزاهمیشه نشان می دهد که این ماده یک کاتالیزور با منشاء بیولوژیکی است - یک آنزیم). قسمت ابتدایی نام است دئوکسی ریبونوکلئاز- نشان می دهد که این آنزیم کدام ترکیب را تجزیه می کند. همه این روش های برش DNA، اول از همه، بر تجزیه و تحلیل دقیق ترکیب آن متمرکز هستند.
مهم ترین اطلاعات موجود در یک مولکول DNA ترتیب تناوب گروه ها است آ, تی, جیو سی، با استفاده از تکنیک های ویژه توسعه یافته به دست می آید. برای این منظور، طیف گسترده ای از آنزیم ها ایجاد شده است که یک توالی کاملاً مشخص را در مولکول DNA پیدا می کنند، به عنوان مثال، سی-تی-جی-سی-آ-جی(و همچنین دنباله مربوطه در زنجیره مقابل جی-آ-سی-جی-تی-سی) و آن را از زنجیره جدا کنید. این خاصیت متعلق به آنزیم Pst I (نام تجاری، از نام آن میکروارگانیسم است. پ rovidencia خیابان uartii که این آنزیم از آن به دست می آید). هنگام استفاده از آنزیم دیگر Pal I، امکان یافتن توالی وجود دارد جی-جی-سی-سی. سپس، نتایج بهدستآمده از عملکرد طیف گستردهای از آنزیمهای مختلف طبق یک طرح از پیش توسعهیافته با هم مقایسه میشوند، در نتیجه میتوان توالی چنین گروههایی را در یک بخش DNA مشخص تعیین کرد. اکنون چنین تکنیک هایی به مرحله استفاده گسترده رسیده اند؛ آنها در زمینه های مختلف به دور از تحقیقات علمی بیوشیمیایی استفاده می شوند، به عنوان مثال، در شناسایی بقایای موجودات زنده یا ایجاد درجه ارتباط.
ساختار RNA
از بسیاری جهات یادآور DNA است، تفاوت این است که در زنجیره اصلی قطعات اسید فسفریک با ریبوز متناوب می شوند و نه با دئوکسی ریبوز (شکل). تفاوت دوم این است که یک هتروسیکل اوراسیل ( U) به جای تیمین ( تی) دیگر هتروسیکل ها آ, جیو سیمانند DNA. اوراسیل با تیمین در غیاب یک گروه متیل متصل به حلقه متفاوت است، در شکل. 10 این گروه متیل با رنگ قرمز مشخص شده است.

برنج. 10. تفاوت تیمین با اوراسیل- عدم وجود یک گروه متیل در ترکیب دوم که با رنگ قرمز در تیمین مشخص شده است.
قطعه ای از یک مولکول RNA در شکل نشان داده شده است. 11، ترتیب گروه بندی ها آ, U, جیو سی، و نسبت کمی آنها ممکن است متفاوت باشد.

شکل 11. قطعه ای از یک مولکول RNA. تفاوت اصلی با DNA وجود گروه های OH در ریبوز (قرمز) و یک قطعه اوراسیل (آبی) است.
زنجیره پلیمری RNA تقریبا ده برابر کوتاهتر از DNA است. یک تفاوت اضافی این است که مولکولهای RNA در مارپیچهای دوتایی متشکل از دو مولکول ترکیب نمیشوند، بلکه معمولاً بهصورت یک مولکول واحد وجود دارند که در برخی مناطق میتوانند قطعات مارپیچ دو رشتهای را با خود تشکیل دهند که با بخشهای خطی متناوب میشوند. در مناطق مارپیچ، برهمکنش جفت ها به همان اندازه در DNA مشاهده می شود. جفت هایی که با پیوندهای هیدروژنی به هم متصل می شوند و یک مارپیچ را تشکیل می دهند ( آ-Uو جی-سی، در مناطقی ظاهر می شود که ترتیب گروه ها برای چنین تعاملی مطلوب است (شکل 12).

برای اکثریت قریب به اتفاق موجودات زنده، محتوای کمی جفت است آ-Uبیشتر از جی-سیدر پستانداران 1.5-1.6 بار، در گیاهان - 1.2 بار. انواع مختلفی از RNA وجود دارد که نقش های متفاوتی در یک موجود زنده دارند.
خواص شیمیایی RNA
شبیه خواص DNA است، با این حال، وجود گروههای OH اضافی در ریبوز و محتوای کمتر (در مقایسه با DNA) در نواحی مارپیچ تثبیتشده، مولکولهای RNA را از نظر شیمیایی آسیبپذیرتر میکند. تحت تأثیر اسیدها یا قلیاها، قطعات اصلی زنجیره پلیمری P(O)-O-CH2 به راحتی هیدرولیز می شوند، گروه ها آ, U, جیو سیراحت تر از بین برود در صورت لزوم به دست آوردن قطعات مونومر (مانند موارد در شکل 9)، در حالی که هتروسیکل های مرتبط شیمیایی را حفظ می کنند، از آنزیم های ظریفی به نام ریبونکولئازها استفاده می شود.
مشارکت DNA و RNA در سنتز پروتئین
- یکی از عملکردهای اصلی اسیدهای نوکلئیک. پروتئین ها مهم ترین اجزای هر موجود زنده هستند. ماهیچه ها، اندام های داخلی، بافت استخوانی، پوست و موی پستانداران شامل پروتئین ها. اینها ترکیبات پلیمری هستند که در یک موجود زنده از اسیدهای آمینه مختلف جمع می شوند. در چنین مجموعه ای، اسیدهای نوکلئیک نقش کنترل کننده ای دارند؛ فرآیند در دو مرحله انجام می شود و در هر یک از آنها عامل تعیین کننده جهت گیری متقابل هتروسیکل های حاوی نیتروژن DNA و RNA است.
وظیفه اصلی DNA ذخیره اطلاعات ثبت شده و ارائه آن در لحظه شروع سنتز پروتئین است. در این راستا، افزایش پایداری شیمیایی DNA در مقایسه با RNA قابل درک است. طبیعت مراقبت کرده است تا اطلاعات اولیه تا حد امکان غیرقابل تعرض باشد.
در مرحله اول، بخشی از مارپیچ دوگانه باز می شود، شاخه های آزاد شده از هم جدا می شوند و به صورت گروهی آ, تی, جیو سی، که مشخص شد در دسترس است، سنتز RNA به نام RNA پیام رسان آغاز می شود، زیرا به عنوان کپی از ماتریس، اطلاعات ثبت شده در بخش DNA آشکار شده را با دقت بازتولید می کند. مقابل گروه آ، متعلق به مولکول DNA، قطعه ای از RNA پیام رسان آینده حاوی گروه وجود دارد. U، تمام گروه های دیگر دقیقاً مطابق با چگونگی این اتفاق در هنگام تشکیل یک مارپیچ دوگانه DNA در مقابل یکدیگر قرار دارند (شکل 13).

طبق این طرح، یک مولکول پلیمری از RNA پیام رسان تشکیل می شود که حاوی چندین هزار واحد مونومر است.
در مرحله دوم، DNA الگو از هسته سلول به فضای دور هسته - سیتوپلاسم حرکت می کند. RNA پیام رسان حاصل با به اصطلاح RNA های انتقالی همراه است که آمینو اسیدهای مختلف را حمل می کنند. هر RNA انتقالی، بارگیری شده با یک اسید آمینه خاص، به یک منطقه کاملاً تعیین شده از RNA پیام رسان نزدیک می شود؛ مکان مورد نظر با استفاده از همان اصل تطابق بین گروهی شناسایی می شود. آ
یک جزئیات مهم این است که تعامل موقت بین پیام رسان و RNA انتقالی تنها در سه گروه رخ می دهد، به عنوان مثال، سه گانه سی-سی-Uاسید ماتریکس، فقط سه گانه مربوطه می تواند مناسب باشد جی-جی-آانتقال RNA، که مطمئناً اسید آمینه گلیسین را با خود حمل می کند (شکل 14). به همین ترتیب برای سه گانه جی-آ-Uفقط یک مجموعه می تواند نزدیک شود سی-U-آ، تنها اسید آمینه لوسین را منتقل می کند. بنابراین، توالی گروه ها در RNA پیام رسان نشان می دهد که اسیدهای آمینه به چه ترتیبی باید ترکیب شوند. علاوه بر این، این سیستم حاوی قوانین تنظیمی اضافی به شکل رمزگذاری شده است؛ برخی از توالیها از سه گروه RNA پیامرسان نشان میدهند که سنتز پروتئین باید در این نقطه متوقف شود، یعنی. مولکول به طول لازم رسیده است.
در شکل نشان داده شده است. 14 سنتز پروتئین با مشارکت یکی دیگر انجام می شود - نوع سوم اسیدهای RNA؛ آنها بخشی از ریبوزوم هستند و بنابراین آنها را ریبوزومی می نامند. ریبوزوم که مجموعهای از پروتئینهای RNA ریبوزومی خاص است، تعامل پیامرسان و RNA انتقالی را تضمین میکند و نقش یک تسمه نقاله را بازی میکند که RNA پیامرسان را یک مرحله پس از اتصال دو اسید آمینه به حرکت در میآورد.
معنای اصلی طرح دو مرحله ای نشان داده شده در شکل. 13 و 14، این است که زنجیره پلیمری یک مولکول پروتئین از اسیدهای آمینه مختلف به ترتیب مورد نظر و دقیقاً مطابق طرحی که به صورت کدگذاری شده روی بخش خاصی از DNA نوشته شده است، جمع می شود. بنابراین، DNA نقطه شروع کل این فرآیند برنامه ریزی شده را نشان می دهد.
در فرآیند زندگی، پروتئین ها به طور مداوم مصرف می شوند، و بنابراین آنها به طور منظم طبق طرح توصیف شده تولید می شوند؛ کل سنتز یک مولکول پروتئین، متشکل از صدها اسید آمینه، در یک موجود زنده در حدود یک دقیقه انجام می شود.
اولین مطالعات در مورد اسیدهای نوکلئیک در نیمه دوم قرن نوزدهم انجام شد، این درک که تمام اطلاعات در مورد یک موجود زنده در DNA رمزگذاری شده است در اواسط قرن بیستم به دست آمد، ساختار مارپیچ دوگانه DNA در 1953 توسط J. Watson و F. Crick بر اساس داده های تجزیه و تحلیل پراش اشعه ایکس، که به عنوان بزرگترین دستاورد علمی قرن 20 شناخته شده است. در اواسط دهه 70 قرن بیستم. روش هایی برای رمزگشایی ساختار دقیق اسیدهای نوکلئیک ظاهر شد و پس از آن، روش هایی برای سنتز هدفمند آنها توسعه یافت. امروزه، همه فرآیندهایی که در موجودات زنده شامل اسیدهای نوکلئیک اتفاق میافتند، واضح نیستند، و امروزه این یکی از حوزههای علمی است که به شدت در حال توسعه است.
میخائیل لویتسکی